-
Kojima M, Hosoda H, Date Y, Nakazato M,
Matsuo H, Kangawa K 1999 Ghrelin is a
growth-hormone-releasing acylated peptide from
stomach. Nature 402:656û660.
-
Wren AM, Small CJ, Ward HL, Murphy KG, Dakin
CL, Taheri S, Kennedy AR, Roberts GH, Morgan DG,
Ghatei MA, Bloom SR 2000 The novel
hypothalamic peptide ghrelin stimulates food intake
and growth hormone secretion. Endocrinology
141:4325û4328.
-
Takaya K, Ariyasu H, Kanamoto N, Iwakura H,
Yoshimoto A, Harada M, Mori K, Komatsu Y, Usui T,
Shimatsu A, Ogawa Y, Hosoda K, Akamizu T, Kojima M,
Kangawa K, Nakao K 2000 Ghrelin strongly
stimulates growth hormone release in humans. J Clin
Endocrinol Metab 85:4908û4911.
-
Tschop M, Smiley DL, Heiman ML 2000
Ghrelin induces adiposity in rodents. Nature
407:908û913.
-
Nakazato M, Murakami N, Date Y, Kojima M,
Matsuo H, Kangawa K, Matsukura S 2001 A
role for ghrelin in the central regulation of
feeding. Nature 409:194û198.
-
Wren AM, Small CJ, Abbott CR, Dhillo WS,
Seal LJ, Cohen MA, Batterham RL, Taheri S, Stanley
SA, Ghatei MA, Bloom SR 2001 Ghrelin causes
hyperphagia and obesity in rats. Diabetes
50:2540û2547.
-
Wren AM, Seal LJ, Cohen MA, Brynes AE, Frost
GS, Murphy KG, Dhillo WS, Ghatei MA, Bloom SR
2001 Ghrelin enhances appetite and increases food
intake in humans. J Clin Endocrinol Metab 86:5992.
-
Schneider LF, Warren MP 2006
Functional hypothalamic amenorrhea is associated
with elevated ghrelin and disordered eating. Fertil
Steril 86:1744û1749.
-
Tanaka M, Tatebe Y, Nakahara T, Yasuhara D,
Sagiyama K, Muranaga T, Ueno H, Nakazato M, Nozoe S,
Naruo T 2003 Eating pattern and the effect
of oral glucose on ghrelin and insulin secretion in
patients with anorexia nervosa. Clin Endocrinol (Oxf)
59:574û579.
-
Misra M, Miller KK, Kuo K, Griffin K, Stewart V,
Hunter E, Herzog DB, Klibanski A 2005
Secretory dynamics of leptin in adolescent girls
with anorexia nervosa and healthy adolescents. Am J
Physiol Endocrinol Metab 289:E373ûE381.
-
Germain N, Galusca B, Le Roux CW, Bossu C, Ghatei
MA, Lang F, Bloom SR, Estour B 2007
Constitutional thinness and lean anorexia nervosa
display opposite concentrations of peptide YY,
glucagon-like peptide 1, ghrelin, and leptin. Am J
Clin Nutr 85:967û971.
-
Katz MG, Vollenhoven B 2000 The
reproductive endocrine consequences of anorexia
nervosa. BJOG 107:707û713.
-
Helmreich DL, Cameron JL 1992 Suppression
of luteinizing hormone secretion during food
restriction in male rhesus monkeys (Macaca
mulatta): failure of naloxone to restore normal
pulsatility. Neuroendocrinology 56:464û473.
-
Whisnant CS, Harrell RJ 2002 Effect of
short-term feed restriction and refeeding on serum
concentrations of leptin, luteinizing hormone and
insulin in ovariectomized gilts. Domest Anim
Endocrinol 22:73û80.
-
Loucks AB, Thuma JR 2003 Luteinizing
hormone pulsatility is disrupted at a threshold of
energy availability in regularly menstruating women.
J Clin Endocrinol Metab 88:297û311.
-
Pohl CR, Richardson DW, Hutchison JS, Germak JA,
Knobil E 1983 Hypophysiotropic signal
frequency and the functioning of the
pituitary-ovarian system in the rhesus monkey.
Endocrinology 112:2076û2080.
-
Vulliemoz NR, Xiao E, Xia-Zhang L, Germond M, Rivier
J, Ferin M 2004 Decrease in luteinizing
hormone pulse frequency during a five-hour
peripheral ghrelin infusion in the ovariectomized
rhesus monkey. J Clin Endocrinol Metab 89:5718û5723.
-
Furuta M, Funabashi T, Kimura F 2001
Intracerebroventricular administration of ghrelin
rapidly suppresses pulsatile luteinizing hormone
secretion in ovariectomized rats. Biochem Biophys
Res Commun 288:780û785.
-
Kluge M, Schussler P, Uhr M, Yassouridis A, Steiger
A 2007 Ghrelin suppresses secretion of
luteinizing hormone in humans. J Clin Endocrinol
Metab 92:3202û3205.
-
Ferin M 2006 Stress and the reproductive
System. In: Neill J, ed. Physiology of reproduction.
3rd ed. San Diego: Elsevier; 2627û2696.
-
Feng YJ, Shalts E, Xia LN, Rivier J, Rivier C, Vale
W, Ferin M 1991 An inhibitory effects of
interleukin-1a on basal gonadotropin release in the
ovariectomized rhesus monkey: reversal by a
corticotropin-releasing factor antagonist.
Endocrinology 128:2077û2082.
-
Shalts E, Feng YJ, Ferin M 1992 Vasopressin
mediates the interleukin-1α-induced decrease in
luteinizing hormone secretion in the ovariectomized
rhesus monkey. Endocrinology 131:153û158.
-
Rivier JE, Kirby DA, Lahrichi SL, Corrigan A, Vale
WW, Rivier CL 1999 Constrained
corticotropin releasing factor antagonists (astressin
analogues) with long duration of action in the rat.
J Med Chem 42:3175û3182
begin_of_the_skype_highlighting 3175û3182 end_of_the_skype_highlighting.
-
Broadbear JH, Winger G, Rivier JE, Rice KC, Woods JH
2004 Corticotropin-releasing hormone antagonists,
astressin B and antalarmin: differing profiles of
activity in rhesus monkeys. Neuropsychopharmacology
29:1112û1121.
-
Xiao E, Xia-Zhang L, Vulliemoz N, Rivier J, Ferin M
2007 Astressin B, a corticotropin-releasing hormone
receptor antagonist, accelerates the return to
normal luteal function after an inflammatory-like
stress challenge in the rhesus monkey. Endocrinology
148:841û848.
-
Xiao E, Xia L, Shanen D, Khabele D, Ferin M
1994 Stimulatory effects of interleukin-induced
activation of the hypothalamo-pituitary-adrenal axis
on gonadotropin secretion in ovariectomized monkeys
replaced with estradiol. Endocrinology
135:2093û2098.
-
Van
Vugt DA, Diefenbach WD, Alston E, Ferin M
1985 Gonadotropin-releasing hormone pulses in third
ventricular cerebrospinal fluid of ovariectomized
rhesus monkeys: correlation with luteinizing hormone
pulses. Endocrinology 117:1550û1558.
-
Rivier CL, Grigoriadis DE, Rivier JE 2003
Role of corticotropin-releasing factor receptors
type 1 and 2 in modulating the rat
adrenocorticotropin response to stressors.
Endocrinology 144:2396û2403.
-
Mozid AM, Tringali G, Forsling ML, Hendricks MS,
Ajodha S, Edwards R, Navarra P, Grossman AB,
Korbonits M 2003 Ghrelin is released from
rat hypothalamic explants and stimulates
corticotrophin-releasing hormone and
arginine-vasopressin. Horm Metab Res 35:455û459.
-
Rivier C, Vale W 1984 Influence of
corticotropin-releasing factor on reproductive
functions in the rat. Endocrinology 114:914û921.
-
Petraglia F, Sutton S, Vale W, Plotsky P
1987 Corticotropin-releasing factor decreases plasma
luteinizing hormone levels in female rats by
inhibiting gonadotropin-releasing hormone release
into hypophysial-portal circulation. Endocrinology
120:1083û1088.
-
Olster DH, Ferin M 1987
Corticotropin-releasing hormone inhibits
gonadotropin secretion in the ovariectomized rhesus
monkey. J Clin Endocrinol Metab 65:262û267.
-
Williams CL, Nishihara M, Thalabard JC, Grosser PM,
Hotchkiss J, Knobil E 1990
Corticotropin-releasing factor and
gonadotropin-releasing hormone pulse generator
activity in the rhesus monkey. Electrophysiological
studies. Neuroendocrinology 52:133û137.
-
Armeanu MC, Berkhout GM, Schoemaker J 1992
Pulsatile luteinizing hormone secretion in
hypothalamic amenorrhea, anorexia nervosa, and
polycystic ovarian disease during naltrexone
treatment. Fertil Steril 57:762û770.
-
Berga SL 1996 Functional hypothalamic
chronic anovulation. In: Adashi EY, Rock JA,
Rosenwaks Z, eds. Reproductive endocrinology,
surgery and technology. Philadelphia:
Lippincott-Raven; 1061û1075.
-
De
Souza MJ, Leidy HJ, OÆDonnell E, Lasley B, Williams
NI 2004 Fasting ghrelin levels in
physically active women: relationship with menstrual
disturbances and metabolic hormones. J Clin
Endocrinol Metab 89:3536û3542
begin_of_the_skype_highlighting 3536û3542 end_of_the_skype_highlighting.
-
Xiao E, Xia-Zhang L, Ferin M 2000
Inhibitory effects of endotoxin on LH secretion in
the ovariectomized monkey are prevented by naloxone
but not by an interleukin-1 receptor antagonist.
Neuroimmunomodulation 7:6û15.
-
Willesen MG, Kristensen P, Romer J 1999
Co-localization of growth hormone secretagogue
receptor and NPY mRNA in the arcuate nucleus of the
rat. Neuroendocrinology 70:306û316.
-
Kamegai J, Tamura H, Shimizu T, Ishii S, Sugihara H,
Wakabayashi I 2001 Chronic central infusion
of ghrelin increases hypothalamic neuropeptide Y and
agouti-related protein mRNA levels and body weight
in rats. Diabetes 50:2438û2443.
-
Hahn TM, Breininger JF, Baskin DG, Schwartz MW
1998 Coexpression of Agrp and NPY in
fasting-activated hypothalamic neurons. Nat Neurosci
1:271û272.
-
Mizuno TM, Makimura H, Silverstein J, Roberts JL,
Lopingco T, Mobbs CV 1999 Fasting regulates
hypothalamic neuropeptide Y, agouti-related peptide,
and proopiomelanocortin in diabetic mice independent
of changes in leptin or insulin. Endocrinology
140:4551û4557.
-
Clark JT, Kalra PS, Crowley WR, Kalra SP
1984 Neuropeptide Y and human pancreatic polypeptide
stimulate feeding behavior in rats. Endocrinology
115:427û429.
-
Larsen PJ, Tang-Christensen M, Stidsen CE, Madsen K,
Smith MS, Cameron JL 1999 Activation of
central neuropeptide Y Y1 receptors potently
stimulates food intake in male rhesus monkeys. J
Clin Endocrinol Metab 84:3781û3791
begin_of_the_skype_highlighting 3781û3791 end_of_the_skype_highlighting.
-
Small CJ, Kim MS, Stanley SA, Mitchell JR, Murphy K,
Morgan DG, Ghatei MA, Bloom SR 2001 Effects
of chronic central nervous system administration of
agouti-related protein in pair-fed animals. Diabetes
50:248û254.
-
Koegler FH, Grove KL, Schiffmacher A, Smith MS,
Cameron JL 2001 Central melanocortin
receptors mediate changes in food intake in the
rhesus macaque. Endocrinology 142:2586û2592.
-
Chen HY, Trumbauer ME, Chen AS, Weingarth DT, Adams
JR, Frazier EG, Shen Z, Marsh DJ, Feighner SD, Guan
XM, Ye Z, Nargund RP, Smith RG, Van der Ploeg LH,
Howard AD, MacNeil DJ, Qian S 2004
Orexigenic action of peripheral ghrelin is mediated
by neuropeptide Y and agouti-related protein.
Endocrinology 145:2607û2612.
-
Kaynard AH, Pau KY, Hess DL, Spies HG 1990
Third-ventricular infusion of neuropeptide Y
suppresses luteinizing hormone secretion in
ovariectomized rhesus macaques. Endocrinology
127:2437û2444.
-
Vulliemoz NR, Xiao E, Xia-Zhang L, Wardlaw SL, Ferin
M 2005 Central infusion of agouti-related
peptide suppresses pulsatile luteinizing hormone
release in the ovariectomized rhesus monkey.
Endocrinology 146:784û789.
-
Dimitrov EL, DeJoseph MR, Brownfield MS, Urban JH
2007 Involvement of neuropeptide Y Y1 receptors in
the regulation of neuroendocrine
corticotropin-releasing hormone neuronal activity.
Endocrinology 148:3666û3673.
-
Haskell-Luevano C, Chen P, Li C, Chang K, Smith MS,
Cameron JL, Cone RD 1999 Characterization
of the neuroanatomical distribution of
agouti-related protein immunoreactivity in the
rhesus monkey and the rat. Endocrinology
140:1408û1415.
-
Li
C, Chen P, Smith MS 2000 Corticotropin
releasing hormone neurons in the paraventricular
nucleus are direct targets for neuropeptide Y
neurons in the arcuate nucleus: an anterograde
tracing study. Brain Res 854:122û129.
-
Martini AC, Fernandez-Fernandez R, Tovar S, Navarro
VM, Vigo E, Vazquez MJ, Davies JS, Thompson NM,
Aguilar E, Pinilla L, Wells T, Dieguez C,
Tena-Sempere M 2006 Comparative analysis of
the effects of ghrelin and unacylated ghrelin on
luteinizing hormone secretion in male rats.
Endocrinology 147:2374û2382.
-
Brundu B, Loucks TL, Adler LJ, Cameron JL, Berga SL
2006 Increased cortisol in the cerebrospinal fluid
of women with functional hypothalamic amenorrhea. J
Clin Endocrinol Metab 91:1561û1565.
-
Berga SL, Mortola JF, Girton L, Suh B, Laughlin G,
Pham P, Yen SS 1989 Neuroendocrine
aberrations in women with functional hypothalamic
amenorrhea. J Clin Endocrinol Metab 68:301û308.
-
Biller BM, Federoff HJ, Koenig JI, Klibanski A
1990 Abnormal cortisol secretion and responses to
corticotropin-releasing hormone in women with
hypothalamic amenorrhea. J Clin Endocrinol Metab
70:311û317.
-
Laughlin GA, Dominguez CE, Yen SS 1998
Nutritional and endocrine-metabolic aberrations in
women with functional hypothalamic amenorrhea. J
Clin Endocrinol Metab 83:25û32.
-
Otto B, Cuntz U, Fruehauf E, Wawarta R, Folwaczny C,
Riepl RL, Heiman ML, Lehnert P, Fichter M, Tschop M
2001 Weight gain decreases elevated plasma ghrelin
concentrations of patients with anorexia nervosa.
Eur J Endocrinol 145:669û673.
-
Williams NI, Helmreich DL, Parfitt DB,
Caston-Balderrama A, Cameron JL 2001
Evidence for a causal role of low energy
availability in the induction of menstrual cycle
disturbances during strenuous exercise training. J
Clin Endocrinol Metab 86:5184û5193.
|
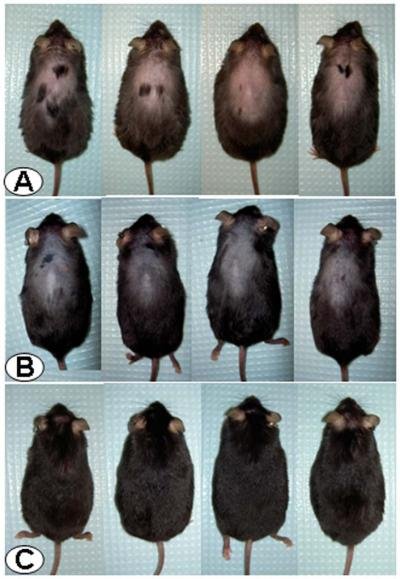 Giunti
al momento della sperimentazione, i ricercatori hanno iniettato per 5 giorni lÆastressina-B
nelle cavie stressate e calve e li hanno quindi rimessi assieme al gruppo di
controllo.
Giunti
al momento della sperimentazione, i ricercatori hanno iniettato per 5 giorni lÆastressina-B
nelle cavie stressate e calve e li hanno quindi rimessi assieme al gruppo di
controllo.